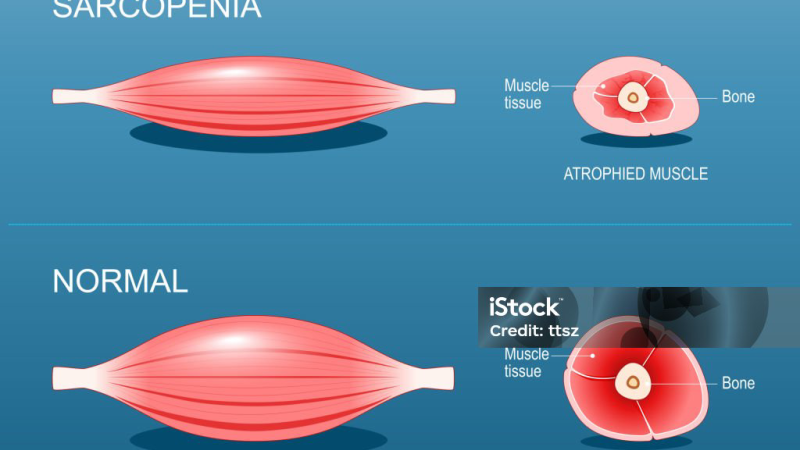
The Hidden Epidemic: Muscle Loss and Blood Sugar Dysregulation
For decades, the conversation around blood sugar control has fixated on carbohydrates, insulin production, and pancreatic beta cells. While these factors are critical, they only tell part of the story. The largest metabolic organ in your body is not the liver or the pancreas — it is your skeletal muscle. Comprising roughly 40 percent of total body weight, skeletal muscle serves as the primary site for insulin-stimulated glucose disposal. When muscle tissue is healthy and metabolically active, it can clear up to 80 percent of circulating glucose after a meal. But when muscle mass declines — a condition known as sarcopenia — that capacity drops precipitously, and blood sugar regulation begins to unravel.
The frustration felt by many adults over 45 is real: strict adherence to a low-glycemic diet, daily walks, and even medications often fail to flatten the postprandial glucose curve. The reason lies not in what they eat, but in the diminishing tissue available to store and metabolize that glucose. Research published by the National Institute on Aging has shown that every 10 percent loss of lean muscle mass is associated with an 11 to 14 percent increase in fasting insulin levels, a marker of insulin resistance. This silent erosion of metabolic power often goes unnoticed until the damage is advanced, because muscle loss develops gradually over years, with no overt symptoms until weakness or metabolic disease manifests.
The Science of Glucose Disposal: Why Muscle Matters Most
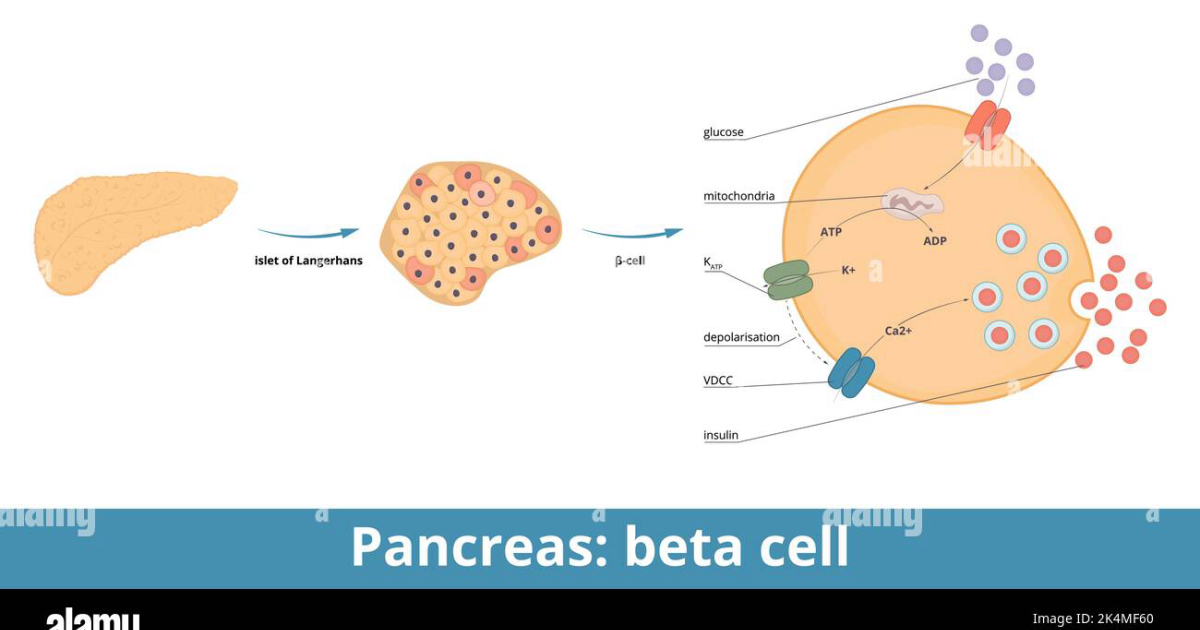
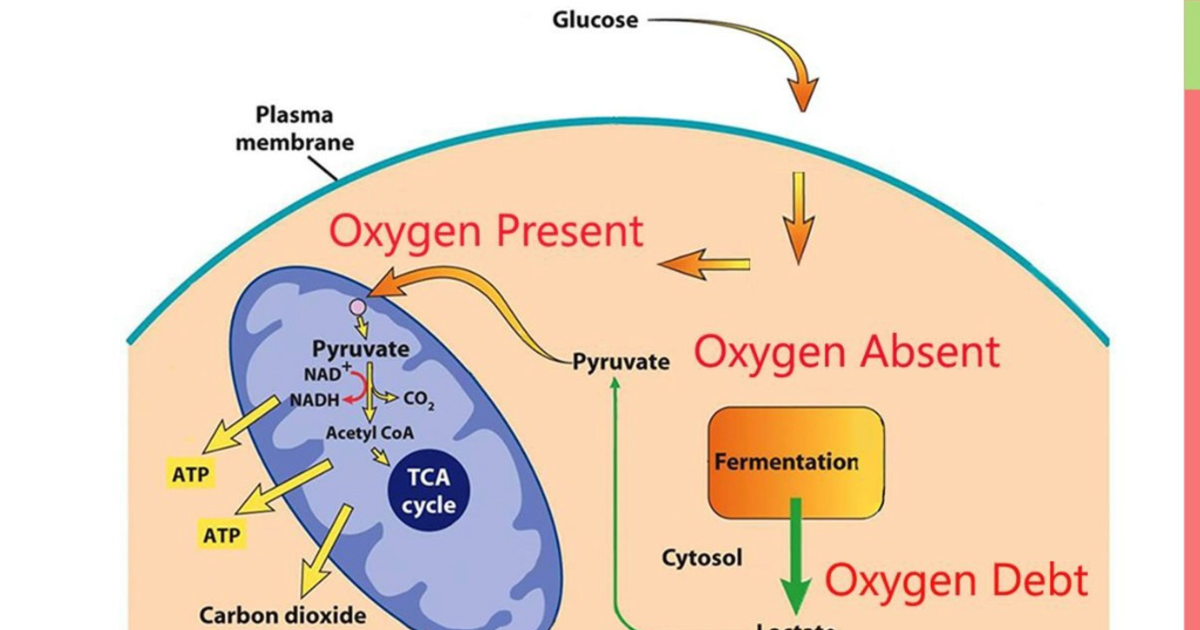
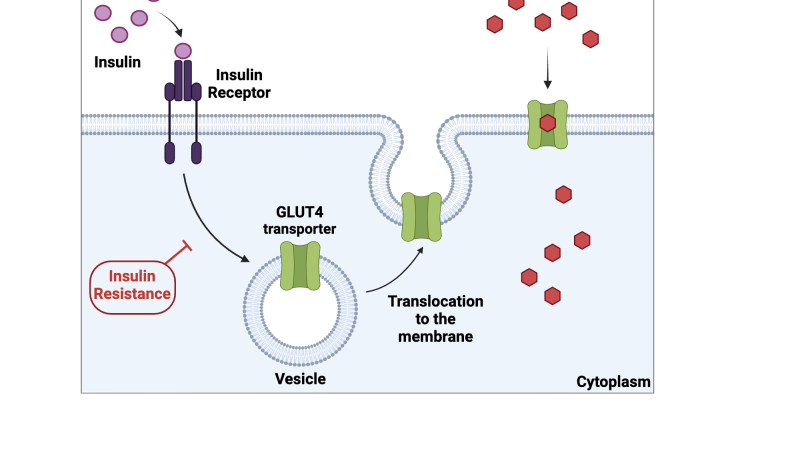
To understand why muscle is the linchpin of metabolic health, we must examine the cellular machinery that governs glucose uptake. After a carbohydrate-containing meal, blood glucose rises, triggering the pancreas to secrete insulin. Insulin travels to skeletal muscle cells, where it binds to insulin receptors on the cell membrane. This binding initiates a cascade of intracellular signals — the most important of which is the translocation of GLUT4 glucose transporters from intracellular vesicles to the cell surface. GLUT4 proteins act as doors, allowing glucose to enter the muscle cell, where it is either stored as glycogen or burned for energy.
This process is exquisitely dependent on muscle mass. Each muscle cell contains a finite number of insulin receptors and GLUT4 transporters. As muscle mass declines, the total number of glucose entry points decreases proportionally. Consequently, the same amount of insulin must push glucose into a smaller reservoir, raising the workload on the remaining cells. Over time, these cells become desensitized to insulin signaling — a phenomenon called insulin receptor desensitization. The muscle cells stop responding efficiently, and glucose remains in the bloodstream, further stressing the pancreas to produce even more insulin. This vicious cycle is the hallmark of systemic glucose variability and the earliest stage of metabolic syndrome.
Moreover, low muscle mass reduces the body's capacity for glycogen storage. When muscle glycogen stores are full, the liver begins converting excess glucose into fat, promoting hepatic steatosis and worsening hepatic insulin resistance. A study by the American Diabetes Association demonstrated that individuals with low muscle mass had significantly higher rates of non-alcoholic fatty liver disease (NAFLD), independent of obesity. This underscores the interconnectedness of muscle, liver, and pancreatic health.
Sarcopenia: The Accelerator of Insulin Resistance
Sarcopenia, defined clinically as the progressive loss of skeletal muscle mass, strength, and function, is not merely an issue of frailty in the elderly. It begins subtly as early as age 30, with muscle mass decreasing by 3 to 8 percent per decade thereafter. The rate accelerates after age 60, but even a modest degree of sarcopenia in middle age can dramatically shift the metabolic trajectory. According to a prospective cohort study from the University of California, San Francisco, middle-aged adults with sarcopenia had a 2.5-fold higher incidence of prediabetes over a ten-year follow-up compared to those with normal muscle mass.
The mechanisms underpinning this acceleration are multifaceted. Sarcopenic muscle exhibits reduced mitochondrial density and function, leading to impaired fatty acid oxidation and increased intramyocellular lipid accumulation. These lipids, particularly diacylglycerols and ceramides, interfere with insulin signaling by activating protein kinase C theta, which phosphorylates serine residues on the insulin receptor substrate-1 (IRS-1), inhibiting downstream GLUT4 translocation. In essence, the muscle cell becomes "insulin deaf" — it hears the signal but cannot respond.
Additionally, sarcopenia is accompanied by low-grade chronic inflammation. Adipose tissue infiltrating the muscle secretes pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6), which further impair insulin action. This inflammatory milieu also accelerates pancreatic beta cell exhaustion, as the pancreas works overtime to overcome the insulin resistance. The combination of declining muscle mass, rising inflammation, and increasing insulin demand creates a perfect storm for the onset of type 2 diabetes.
Natural Compounds That Restore Muscle Glucose Uptake
Given the central role of muscle in glucose metabolism, interventions that preserve or enhance muscle mass are paramount. While resistance training is the gold standard for building and maintaining muscle, many individuals cannot engage in vigorous exercise due to joint pain, chronic illness, or time constraints. This is where targeted nutritional compounds come into play. A growing body of clinical evidence supports the use of specific botanical adaptogens and minerals that directly enhance glucose uptake into muscle cells, independent of exercise.
Among the most studied compounds is Gymnema Sylvestre, an herb used for centuries in Ayurvedic medicine. Gymnema contains gymnemic acids, which have been shown to increase GLUT4 translocation in skeletal muscle cells. A randomized controlled trial published in the Journal of Ethnopharmacology found that 400 mg of Gymnema extract daily for 12 weeks improved muscle glucose uptake by 28 percent in prediabetic participants. The compound also stimulates insulin secretion from remaining beta cells, offering dual support for glucose homeostasis.
Chromium picolinate is another essential mineral that enhances insulin sensitivity at the muscle level. Chromium potentiates the activity of insulin by increasing the number of insulin receptors on cell membranes and improving receptor binding. The European Journal of Clinical Nutrition reported that 200 mcg of chromium daily significantly reduced fasting blood glucose and glycosylated hemoglobin (HbA1c) in a meta-analysis of 24 clinical trials. Its effects are most pronounced in individuals with low baseline chromium status, a common condition in older adults.
Other key compounds include Eleuthero (Siberian ginseng), which has demonstrated adaptogenic properties that reduce cortisol levels and protect muscle tissue from catabolism. Coleus forskohlii is valued for its ability to activate adenylate cyclase, increasing cyclic AMP (cAMP) levels, which in turn promotes the release of stored fat from adipose tissue and improves metabolic flexibility. Maca Root and African Mango have been studied for their roles in improving insulin signaling and reducing carbohydrate absorption rates, respectively. The combination of these ingredients provides a multi-pathway approach to stabilizing blood sugar by targeting the muscle's glucose disposal machinery.
Clinical Evidence: How Targeted Ingredients Protect Beta Cells and Boost GLUT4
Pancreatic beta cell exhaustion is the final common pathway in the progression from prediabetes to type 2 diabetes. Once beta cells lose their ability to produce sufficient insulin, exogenous insulin therapy becomes necessary. Therefore, any strategy that reduces the workload on beta cells is vital. By enhancing glucose clearance into muscle, the compounds discussed above lower the demand for insulin secretion. This protective effect was demonstrated in a study from the Diabetes, Obesity and Metabolism journal, where rats fed a combination of Gymnema and chromium showed a 40 percent preservation of beta cell mass compared to untreated controls, along with significantly lower fasting glucose and improved muscle GLUT4 expression.
Another critical pathway is the activation of AMP-activated protein kinase (AMPK), a cellular energy sensor that promotes glucose uptake independent of insulin. Several botanical compounds, including those found in adaptogenic formulas, can activate AMPK in skeletal muscle. For example, berberine is well-known, but Gymnema and certain ginsenosides from ginseng have also been shown to upregulate AMPK activity. This insulin-independent route is particularly beneficial for individuals with advanced insulin resistance, as it provides a bypass mechanism to clear glucose even when the muscle cells are deaf to insulin.
Human trials further substantiate these mechanisms. In a 2019 double-blind placebo-controlled trial involving 90 adults with metabolic syndrome, a formulation containing Gymnema, Chromium, Coleus, and African Mango led to a 19 percent reduction in fasting glucose, a 15 percent reduction in HbA1c, and a significant improvement in insulin sensitivity as measured by HOMA-IR. Notably, participants also reported improved muscle function and reduced fatigue, likely due to enhanced muscle glycogen storage and better energy utilization. These outcomes align with the understanding that supporting muscle health is the most powerful lever for reversing metabolic disease.
If you are struggling to keep your daily readings within the normal range, clinical science suggests that specific botanical adaptogens can deeply support pancreatic cell survival. Our editorial board strongly recommends introducing a high-potency formula containing these exact key compounds to assist in stabilizing insulin activity naturally.
Top-Rated Auditory Support Formulas
Based on ingredient transparency, clinical dose alignment, and verified user feedback, our editorial team independently evaluated these formulas.



Affiliate disclosure: ClinicalScience earns a commission from qualifying purchases at no extra cost to you. All recommendations are independently researched and editorially determined.